Del 1.
”I sin tidigare version framställdes kromosomerna som klaviaturen till ett enormt piano med miljontals tangenter… En muterad gen är en tangent som har tappat stämningen. Vid vissa tillfällen – uppmanade man oss att tro – när en hel massa tangenter har tappat stämningen riktigt mycket, blir resultatet en underbar ny melodi: ett kräldjur som förvandlas till en fågel eller en apa förvandlad till en människa. Någonstans på vägen har teorin uppenbarligen spårat ur.” (ur ”Janus” av Arthur Koestler).
Jorden anses idag vara ungefär 4 500 miljoner år gammal. Från livets uppkomst förekom under lång tid endast encelliga organismer, s. k. prokaryoter, men rester av alger har påträffats i så tidiga avlagringar som för 3 500 miljoner sedan i Australien. De första fossilerna från djur dateras till kambrium för 600 miljoner år sedan. Alla olika klasser av djur uppträder därefter stegvis på jorden med perioder om 50 – 100 miljoner år emellan. Först blötdjur (mollusker, ostron och sniglar), därefter fiskar, amfibier (t.ex.grodor), reptiler, däggdjur och fåglar (tab. 1).
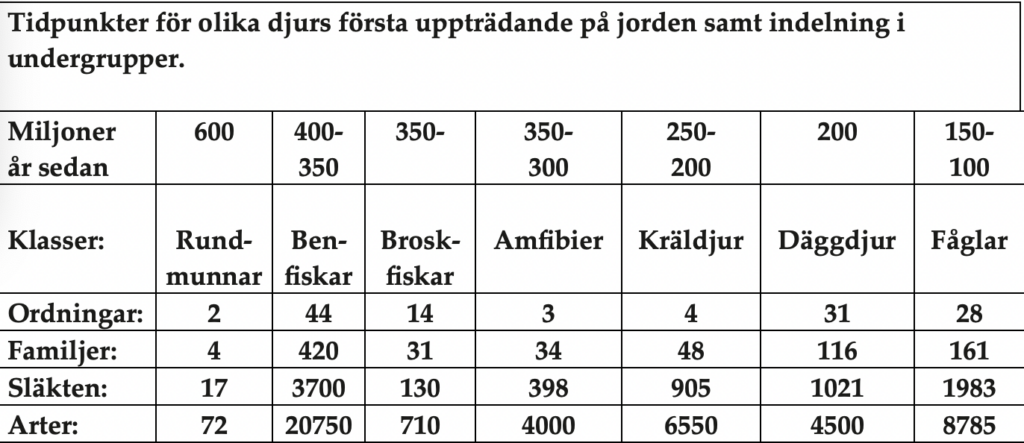
Tabell 1. Den typologiska hierarkiska klassifikationen i arter, släkten, familjer, ordningar och klasser. Siffrorna hämtade och sammanställda från ”World of animals”. Svenska upplagan ”Jordens djur” utgiven av Bonnier Fakta 1986. Svensk huvudgranskare: Prof. Staffan Ulfstrand, Uppsala Universitet. Observera att den absolut talrikaste djurgruppen, insekterna, inte finns med i denna tabell.
Stora gap mellan klasserna – diskontinuitet
Förutom sin likhet i principiell skelettuppbyggnad (homologi) skiljer sig de olika klasserna mycket från varandra. Däggdjuren har hårbeklädnad, svett-körtlar, bröstmjölk, hjärta med fyra kammare, avskilda njurar, stor sex-lagers hjärnbark, diafragma och tre specialiserade öronben. Fåglarna har fjädrar, vingar, genomgångslunga, luftsäckar på olika delar av kroppen, enormt förstorad lill-hjärna etc, etc.
Dessa speciella egenheter finns hos alla arter, släkten, familjer och ordningar inom klassen men inte hos någon art utanför klassen. Därvidlag är alla arter inom klassen lika representativa, och ingen art kan sägas ligga närmare varken mitten eller periferin. Detta var känt som ”typologi” bland dåtidens vetenskapsmän, och detta stora ”gap” mellan de olika klasserna är så uppenbart, att varje litet barn spontant känner igen skillnaden mellan fågel och fisk.
Darwin pläderade för kontinuitet
När Charles Darwin 1831 anträdde sin 5 år långa och numera världsberömda resa till Galapagos-öarna, var han 22 år. Med sig på resan hade han en året innan utgiven bok, ”The Principles of Geology” av Charles Lyell, som visade på jordens långsamma förändring under flera miljarder år. Vid mötet med djurlivet på Galapagosöarna slogs Darwin omedelbart av att fåglar på olika isolerade öar var mera olika varandra än vad han tidigare upplevt, vilket alltså innebar, att arterna inte kunde vara ”oföränderliga”, som man tidigare trott. I betänkande av geografiska klimatskillnader, avstånd mellan öarna osv. spekulerade han över, om ”skapelseberättelsen” verkligen var hållbar. På något sätt, trodde han, måste dessa fågelarter ha utvecklats från en gemensam utgångsart, nu utdöd. Det kunde ha skett som en mycket långsam, kontinuerlig process med små, små steg i likhet med de av Lyell påvisade geologiska förändringarna – men mekanismen bakom förändringen förbryllade honom.
1838 läste han så boken ”Befolkningsfrågan”, som utkommit tidigare, i vilken Thomas Malthus drev tesen, att det alltid föds ett överskott av individer. Ett stort antal av dessa går dock under, medan de mest ”livsdugliga” har störst chans att överleva.
Efter att Darwin läst denna bok, började bitarna falla på plats, och 1842 skrev han ett första 35-sidors utkast på sin teori, som innebar, att det hade funnits förbindelselänkar mellan de olika klasserna, men att dessa förbindelselänkar dött ut och försvunnit. Livet, enligt Darwin, hade börjat spontant med bildning av viktiga molekyler, sedan enstaka celler, växter och djur. Dessa senare utvecklades ur varandra; först fiskar, därefter amfibier, reptiler, fåglar och däggdjur.
Det skulle dock dröja ända till 1859, innan han vågade sig på att offentligt publicera sina idéer i boken ”Om arternas uppkomst”. Anledningen är uppenbar: Teorin hade en rent spekulativ karaktär utan ett enda bevis. Darwin behövde helt enkelt tid på sig för att samla ”indirekta bevis” och att argumentera kring dessa.
Främsta kritiken från vetenskapsmännen
Boken blev genast en best-seller bland intellektuella. Den blev dock inte lika populär hos den tidens mest kända vetenskapsmän Georges Cuvier, Louis Agassiz, Richard Owen och Charles Lyell m. fl, som påtalade bristen på ”bevis”. Kritikerna var inte direkt fientliga till tanken på en kontinuerlig, långsam process ”från den lägsta till högsta formen av organism”, men ”varför”, som en ledande paleontolog, François Jules Pictet, 1860 uttryckte det: ”Varför finner vi inte dessa små, gradvisa förändringar bland de fossila fynden, och varför hittar vi inte mera mellanformer, istället för att samla tusentals identiska fynd ….Varför och enligt vilka konstiga sannolikhetsregler händer det, att de arter, som vi vanligen och i mest överflöd hittar i nyligen upptäckta områden, i den absolut överväldigande majoriteten av fallen är arter, som vi redan har i våra samlingar?”
Darwins erkände bristen på bevis. För att teorin skulle kunna accepteras, borde man kräva, att bland de fossila resterna finna ”… ett orimligt stort antal (’hundratusentals’) övergångsformer…” mellan de olika klasserna, sade han – och försvarade sig sedan med, att endast en liten del av jordytan ännu utforskats.
Inga ”mellanformer” funna
Idag, när praktiskt taget hela jordytan samt havet utforskats, och antalet paleontologer utökats från en handfull till tiotusentals, är strävandena lika resultatlösa. 99% av alla fossila fynd anses ha insamlats efter Darwins död. Trots fossila fynd av 100 000 olika arter, där de s. k. övergångsformerna enligt Darwin borde räknas i ’hundratusentals’, har ännu inga mellanformer mellan de olika klasserna återfunnits.
Lynn Margulis, biologiprofessor vid universitetet i Massachusetts, berättade, att hon en gång under sina många konferenstal frågat, om molekylärbiologerna i auditoriet kunde nämna något enda otvetydigt exempel på bildning av en ny art genom ackumulering av mutationer. Tystnad. Ingen kunde naturligtvis möta upp till detta krav.
Eller som Stanley (1979) uttryckte det: ”…den fossila vittnesbörden misslyckas med att visa ett enda exempel på gradvis evolution, som ger en stor morfologisk övergång…”.
Detta är ”paleontologins yrkeshemlighet”, enligt Stephen Jay Gould, som företräder en särskild riktning inom evolutionsbiologin (”Theory of Punctuated Equilibrium”). Det beror dock inte på otillräckligt letande. Av nutida landgående ryggradsdjur i världen har 79,1% återfunnits som fossila rester (87,8% om fåglar undantas). I Nordamerika, anser G. G. Simpson, är uppteckningen av dessa djur helt komplett. Det finns – efter mer än hundrafemtio års idogt letande – endast en handfull arter, som man anser skulle kunna vara ’övergångsformer’.
Man har t.ex. talat om lungfisken som en mellanform mellan fisk och amfibie – men lungfisken har två separata och fullt utvecklade system för andning, gälar och lungor, och kan ligga nergrävd i dyn under torrperioder i upp till 3 år. Lungan i detta fall är ingen ’mellanform’ under utveckling.
En form av kvastfeningen (Rhipidistia) ansågs länge vara en mellanform mellan fisk och amfibie. Denna troddes vara utdöd sedan flera hundra miljoner år, men 1938 hittades en levande individ i Indiska Oceanen. En undersökning visade, att kvastfeningen var helt och hållet fisk, när man såg till mjukdelar och fysiologi.
Seymouria (’pansargroddjur’) ansågs vara ett mellanting mellan amfibie och reptil, men reproduktionssystemet var helt och hållet amfibiskt osv, osv.
Det finns således inget fynd av någon otvetydig ’mellanform’. Det är dessutom väldigt subjektivt att utifrån grov morfologi avgöra tillhörighet, eftersom biologin till 99% bestäms av mjukdelarna. Hur skiljs t.ex. pungdjur från livmoderdjur, som i de flesta fall är praktiskt taget identiska, vad gäller skelettmorfologi?
Variation är en sak – evolution är en annan
Det skulle dröja nästan hundra år, innan ett enda exempel av ”naturligt urval” kunde pekas ut – Kettlewells prickiga mal från tidigt 1950-tal. Då hade teorin redan för många år sedan omvandlats till en dogm. Därefter har man faktiskt kunnat visa på flera exempel på ”naturligt urval” och även uppkomst av nya arter. Man har t.ex. på Hawaii kunnat följa utvecklingen av 600 – 700 unika arter av bananflugan från en eller två ursprungskolonier. Där finns också 4 300 unika insektsarter, som man med säkerhet vet har utvecklats från endast 250 ursprungskolonier. Flera exempel finns från andra arter och andra delar av världen, t.ex. stortrutar och fiskmåsar i Sibirien.
Så Darwins ”speciella teori” är riktig, och ”mikroevolution” är möjlig, men det rör sig om triviala förändringar. Trots genetikernas tålmodiga arbete med tusentals generationer av bananflugan, har detta inte frambringat några evolutionära förbättringar, och bananflugorna har aldrig blivit något annat än bananflugor. Det finns inga exempel på, att en art förändrat sig, så att den nu tillhör ett nytt släkte, familj, ordning eller klass. Det finns alltså inga som helst tecken till evolution (i bemärkelsen ”från lägre till högre”) i naturen.
Sedan antiken har varit känt, att man vid avelsförsök når ett visst tak. Fossiler av arter, som hittats under senare tid, ser likadana ut som fossiler av samma art från den tid, då den första gången uppträdde på jorden.
Del 2.
Också proteinanalyserna talar emot evolutionsteorin
Sedan 1960 har det varit möjligt att kartlägga proteiner, aminosyra för aminosyra, och således kvantifiera, hur mycket ett protein skiljer sig från ett annat. Sådana analyser har därför gjorts för att på ett objektivt sätt kunna kartlägga släktskap och evolutionära utvecklingsvägar. Om ett släktskap finns, borde den procentuella skillnaden av enskilda aminosyror öka, ju längre från ursprunget (encelliga organismer) man kommer. Dvs. avståndet mellan människa och bakterie borde vara större än avståndet mellan fisk och bakterie.
Cytokromer finns i mitokondrierna hos alla organismer från encelliga bakterier till människa och hemoglobin finns hos alla djur (förutom insekter) och är därför utmärkta för analys. I Margaret Dayhoffs ”Atlas of protein sequence and structure” gjordes 1.089 jämförelser mellan bl. a. cytokrom C och hemoglobin från olika arter. Vad gäller cytokrom C, befinner sig ingen svamp-, växt- eller djurart närmare eller längre från bakterier (64–69%), insekter (27–30%) eller fiskar (13–14%) än någon annan (se fig.2 a-c i slutet av artikeln). När man övergår till aminosyresekvensen i hemoglobin, skiljer sig samtliga djur från blötdjur med 85–87% och från ål med 73–81%. Inga av de högre djuren kan sägas ligga närmare eller längre från blötdjur och rundmunnar (ursprungliga fiskar) än de övriga (se fig.2 d-e i slutet av artikeln).
Liknande förhållanden finner man vid övriga proteinanalyser. Med denna mera objektiva och kvantifierbara metod kan den evolutionära utvecklingskedja bakterier – rundmunnar – fiskar – amfibier – reptiler – fåglar – däggdjur, som darwinisterna förfäktar, alltså inte alls styrkas. Tvärtom talar analyserna emot detta. (Se figurerna i slutet av artikeln.)
Apan vid skrivmaskinen
Men Darwin spekulerade inte bara om, huruvida amfibierna utvecklats från fiskar, reptiler från amfibier osv, osv. Det är idag axiomatiskt att uppfatta darwinismen som en teori, som förklarar hela livets uppkomst. ”Det är inte orimligt att anta, att livet uppkom i en vattnig ’soppa’ av förbiologiska organiska föreningar, och att levande organismer senare upp stod genom omgivande kvantiteter av dessa föreningar med hjälp av membran som förvandlade dem till ’celler’. Detta betraktas allmänt som början på den organiska (”darwinistiska”) evolutionen”, heter det t. ex. i The New Encyclopaedia Britannica, och ungefär så beskrivs det i våra biologiböcker.
Entusiasmen för en ’naturlig’ lösning på livets gåta har alltid varit mycket stor, och ingenting tycks vara lättare än att tänka sig en molekyl, som kan regenerera sig själv och därefter omge sig med ett membran fullt med receptorer för kommunikation med omvärlden. Émile Borel utvecklade t. ex. 1913 ett speciellt teorem (fig.1) som, i analogi med evolutionsteorin, slog fast att visst kunde apor skriva Shakespeare genom att slumpvis trycka på tangenterna – bara de fick ”oändligt” lång tid på sig. Ungefär som att man kan bygga slott genom att under oändligt lång tid slumpvis slänga tegelstenar i en hög.
Hur förhåller det sig med sanningshalten i dessa påståenden? Jorden är ju inte oändligt gammal utan endast 4 500 miljoner år. Låt oss ta de språkliga möjligheterna på en skrivmaskin som exempel:
Inom engelska språket finns 500 ord på 3 bokstäver (t.ex. ”hat”, ”cat” etc.), men kombinationsmöjligheten är 263 (≈17 000). Endast ett av 34 av på detta sätt slumpvis bildade ord skulle således ha någon mening. Resten är nonsens.
Vidare i jämförelsen finns 10 000 och 1000 ord på 7 respektive 12 bokstäver (t.ex. ”English”, ”require”, ”construction”, ”reasonable”) men 267 (≈ 8 x 109) resp. 2612 (≈1017) kombinationsmöjligheter. När man räknar på detta så framgår att endast 1/100 000 respektive 1/1014 är här meningsfulla ord. Resten är nonsens.
1014 är ett mycket stort tal. Om man skulle tänka sig en apa, som slumpvis tryckte på en skrivmaskins tangenter, ungefär en bokstav var femte sekund, skulle det ta 1 000 miljoner år att slumpvis kunna bilda ett enda meningsfullt 12-bokstäversord.
Men litterära verk består ju inte av enstaka ord utan av meningsfulla meningar. Lingvister har uppskattat antalet meningsfulla 100-bokstavsmeningar i engelska språket till 1025, men kombinationsmöjligheterna av 100 bokstäver är avsevärt mycket större, 26100 (=10130). Detta innebär, att man av en slump bara skulle kunna konstruera 1 meningsfull mening per 10105 nonsensmeningar.
Analogin med apan var alltså felaktig. Nu börjar vi få en känsla för möjligheten att kunna skapa meningsfull DNA-information av en ren slump.

Figur 1. Utifrån ett matematiskt teorem skapade Émile Borel 1913 det s.k. apteoremet som sedan flitigt har citerats som stöd för evolutionsteorin. Det ’oändliga apteoremet’ slår fast att ”en apa som slumpvis trycker på skrivmaskinstangenterna för en oändligt lång tid kommer sannolikt att slutligen ha skrivit vilken särskilt utvald text som helst, som samtliga böcker i Frankrikes Nation-albibliotek…eller William Shakespeares samlade verk.”
”Slumpen” + ”naturligt urval” förklarar ej livets uppkomst
I diskussionen om, hur livet uppstod, anses proteinet ha en särställning, eftersom det är självreplikerande via ribosomerna, som också består av ett 50-tal proteiner. Ett protein består i allmänhet av 100 – 500 aminosyror. I analogi med den förra diskussionen om språket blir då antalet möjliga proteinmolekyler, som kan bildas inom denna ram med 20 olika aminosyror (primärstrukturen) = 20100+20101+20102+…+20498+20499+ 20500 ≈10780. Samtidigt uppskattas antalet olika aktiva proteinmolekyler i kroppen till 100.000. Man kan räkna ut att sannolikheten för att av en slump bilda en enda sådan aktiv proteinmolekyl är mindre än 1 chans på 10775, och sannolikheten att av en slump skapa en cell, som samtidigt kan producera alla de 100.000 olika proteinerna i en cell är mindre än 1 chans på 1077.500.000.
Men alla delar av ett protein är kanske inte lika viktiga? Enzymets ”active site” bestående av 16 aminosyror anses t. ex. vara viktigare än övriga delar av molekylen. Astronomerna Sir Fred Hoyle och Chandra Wickramasinghe har angivit sannolikheten för att slumpvis bilda en sådan liten unik aminosyresekvens (16 aminosyror) av ett enzym till 1/2016≈ 1/1020. Med cellens beräknade 2.000 enzymer blir sannolikheten att slumpvis bilda en cell med dessa ”active sites” ändå så liten som 1/1040.000. Inte ens med dessa siffror skulle en cell kunna bildas av en slump, om så hela universum bestod av ”organisk ursoppa”. Därmed är ingenting ännu sagt om genialiteten i cellens uppbyggnad. Att märka är också, att alla atomer i universum beräknats till 1070.
Strikta regler vid vetenskapliga undersökningar
Vid vetenskapliga undersökningar vill vi veta, i vilken mån resultatet av undersökningen kan förklaras av en slumpvariation. Kan en sådan uteslutas (sannolikheten är mindre än 5%), förklaras resultatet av andra faktorer än slumpen. För att få kontroll över tänkbara faktorers inverkan på resultatet ’randomiseras’ patienterna avseende kön, ålder etc, etc, och undersökningen ’blindas’ eller ’dubbelblindas’ för att få kontroll på psykiska faktorers inverkan, så att allting är lika förutom den undersökta faktorns inverkan (medicin eller åtgärd). Allt detta är välkänt för alla, som sysslar med vetenskap. Om sannolikheten för slumpen är mindre än eller lika med 1/20, är oddsen för andra faktorers inverkan gentemot slumpen 19 mot 1.
Tillämpas detta på våra exempel med bildningen av protein ses, att oddsen, för att andra faktorer än slumpen spelat roll, är ungefär 10510 mot 1 och för bildningen av celler 1040.000 mot 1 eller 1077.500.000 mot 1.
Med detta som bakgrund framstår Jaques Monods uttalande (Nobelpriset 1965) som minst sagt djärvt, då han jämför utvecklingen med ett ”gigantiskt lotteri” och konkluderar att: ”Endast slumpen ligger till grund för varje innovation, varje skapelse i biosfären, den rena slumpen, absolut fri men blind … Denna centrala tes i den moderna biologin är inte längre en bland flera tänkbara hypoteser. Den är idag den enda tänkbara hypotesen, … Och ingenting motiverar antagandet – eller hoppet – att vår uppfattning på denna punkt någonsin kommer att ändras…” Monod, J. ”Slump och nödvändighet” (Stockholm 1972 sid. 25).
Inför evolutionsteorin kollapsar alla vetenskapliga regler
”Dagdrömmar är tillåtna, men vetenskapen skall inte ge efter för dem”, skrev Professor Pierre Grassé, Sorbonne, en gång angående evolutionsteorin. För mig är det ganska självklart, att livets uppkomst inte kan förklaras utifrån slumpmässiga faktorer. Man måste räkna med andra, okända faktorer, och jag blir alltid förvånad, när jag träffar vetenskapligt utbildade människor, som inte inser detta. För dem tycks det inte spela någon roll, om oddsen för andra faktorer mot slumpen är 1040.000 mot 1 eller 1077.500.000 mot 1. Övertygelsen står fast – slumpen och endast slumpen har orsakat detta, eftersom det rör sig om så ”långa tidrymder”.
Det verkar, som om alla vetenskapliga regler kollapsar inför evolutionsteorin, som om det rörde sig om två olika vetenskapliga världar. I den ena världen, som omfattar alla ”bevisande” vetenskaper förutom evolutionsteorin, gäller väldigt strikta vetenskapliga regler; Om oddsen för andra faktorer gentemot slumpen är större än 19:1 gäller, att slumpen är utesluten som orsak till variationerna.
Den andra världen omfattar bara evolutionsteorin. I den världen gäller, att slumpen alltid är orsaken, hur liten sannolikheten för denna än är. Varför har man andra och diametralt motsatta vetenskapliga regler avseende sannolikheter när det gäller evolutionsteorin ?
Proteinanalyser: Om ett släktskap finns, borde den procentuella skillnaden av enskilda aminosyror öka, ju längre från ursprunget (encelliga organismer) man kommer. Dvs. avståndet mellan människa och bakterie borde vara större än avståndet mellan fisk och bakterie.
Figur 2 a-e:
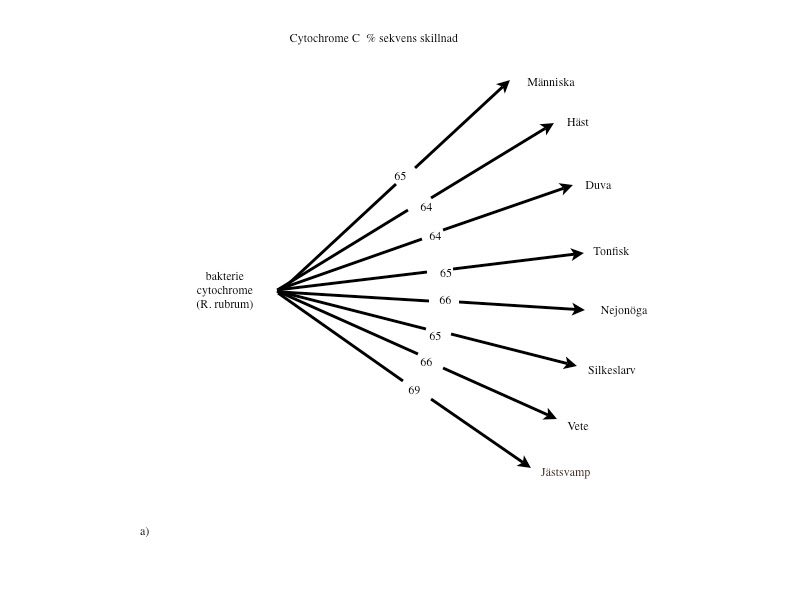
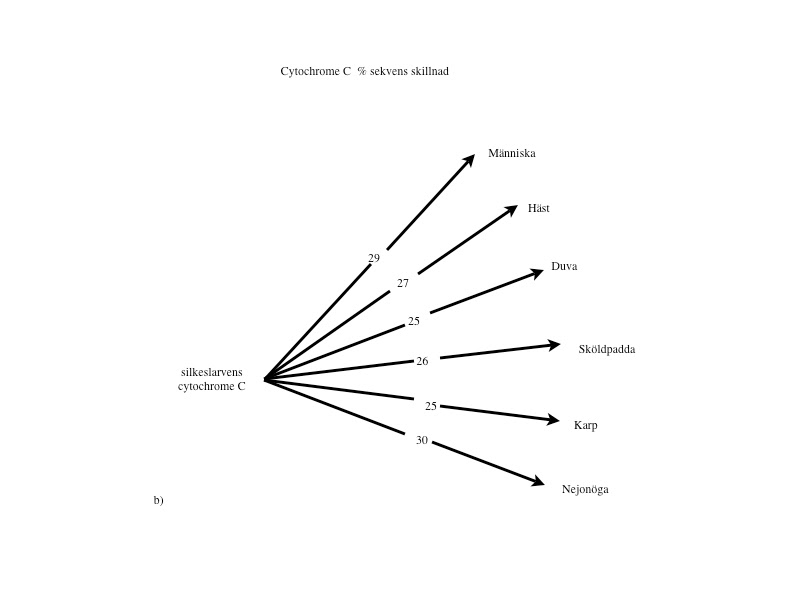
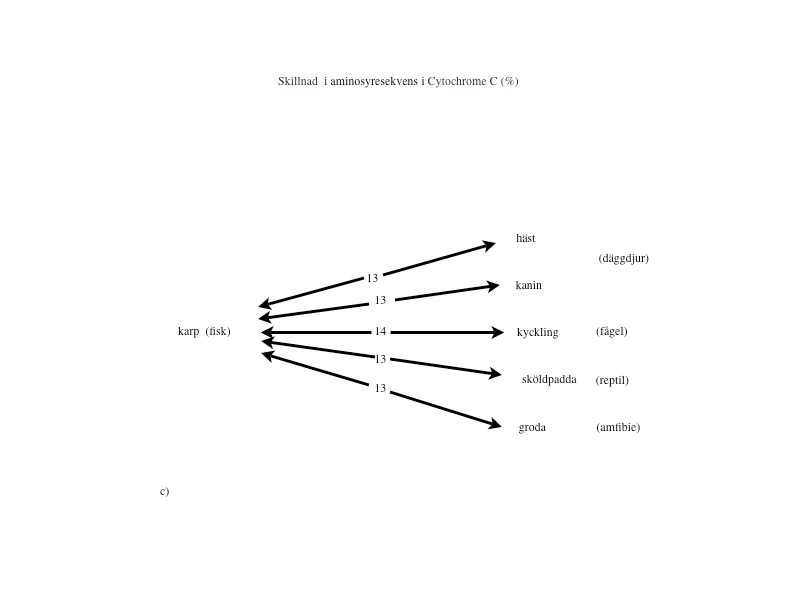


Figur 2 a-e visar ett urval av de 1.089 jämförelser avseende Cytochrome C och Hemoglogin som återfinns i Dayhoffs ”Atlas of protein sequence and structure”. Bilderna är tagna från Michael Dentons bok ’Evolution: A theory in crisis’ och återges i något modifierad form. De visar att vilken klass man väljer att utgå ifrån så kommer alla övriga arter inom de övriga klasserna att befinna sig på lika avstånd. Ingen art inom dessa klasser kan sägas var mera primitiv eller mera utvecklad än de övriga.
Referenser:
1) Arthur Koestler, ’Janus’ (kap. 9-11)
2) Charles Darwin, ’Om arternas uppkomst’
3) Michael Denton (1986): ’Evolution: A theory in crisis’.
4) François Jules Pictet, återgivet i ’Evolution: A theory in crisis’, sid. 104.
5) Lynn Margulis, återgivet i Michael J Behe: ’Darwin’s black box’, sid.26.
6) Stanley, S. (1979) ’Macroevolution’, W.H. Freeman and Co, San Fransisco,
sid. 39.
7) Gould, S. J. (1978) Nov. 26 The Guardian Weekly, vol 119, no 22, sid. 1.
8) ’Evolution’, i The New Encyclopaedia Britannica, vol 7, s 23 och 17 (1975)
9) Margaret Dayhoff, ’Atlas of protein sequence and structure’
10)Émile Borel (1913). ’Mécanique Statistique et Irréversibilité’. J. Phys.
5e série 3 sid. 189–196.
11)Monod, J. ’Slump och nödvändighet’ (Stockholm 1972 sid. 25).
(Denna artikel skrevs av mig första gången 2004 för min facktidskrift ’SFAI-tidningen’. Den återges här i något modifierad form.)